Neurons do not stably participate in the representation of tasks and behaviors. Called representational drift, this calls for new thinking in how the brain represents information.
Localization of Function and Stable Engrams
Ever since Brodmann carved up the cortex into functional domains in the 1800s, the idea of localized function has dominated neuroscientific research and thinking. The general assumption is that activity of a particular area would therefore indicate that the brain is involved in a particular kind of function. Yet this framework has a number of conundrums.
At a macro level, of course sensory information is delivered to specific areas of the brain – there are general architectural principles such that the wire and signals carrying visual information from the eyes end in the visual cortex, the ears in the auditory cortex and so on. It would therefore stand to reason that these areas where information arrives are where it is processed. And broadly studies appear to line up. Generally, which is to say, more often than not, areas involved with visual tasks tend to get more active when there is a visual task at hand and so on.
At the neuronal level the theory that neuroscientists have hung their hat on is one proposed in the early 1940s by Donald Hebb which is the idea of an engram – a group of neurons that together form a representation of a perception or behavior. This paradigm assumes that once formed an engram would be a stable representation, participating neurons faithfully performing their bit. Yet even as this theory seeped into the neuroscientific consciousness, there were already doubts. Hebb’s contemporary, Neurosurgeon Wilder Penfield had been stimulating brains of his patients to map somatosensory and motor function and observed that individuals differed a fair bit, and that in those who had repeated surgeries the representations moved and were not stable.
See related post Stimulation, Sensation and Localization in the Cortex
Yet this work was largely ignored by the neuroscience community, perhaps because it was inconvenient (there was no replacement theory to account for these differences and moving around of representations) or perhaps because fields even as close together as neurosurgery (which looks at humans at a more macro resolution) and neuroscience (which is dominated by cellular investigation in rodents) developed quite separately from one another. Eventually however, the cellular investigations got more sophisticated in being able to track more neurons and track them over time and came to a similar conclusion. That representations or responses of neurons to stimulus and behaviors shift over time – termed representational drift.
Representational Drift in Neurons
The first papers of representational drift in the rodent neuroscience community were published less than ten years ago showing that the coding of spatial location by neurons (‘place codes’) in the CA1 region of the hippocampus were not stable. Not only were the place codes not stable but the dendritic spines that were required to form stable networks were not so stable themselves showing that anatomically networks were reconfiguring. In fact, spatial representations in the hippocampus identified on one day would become so uncorrelated that that spatial decoding accuracy fell to chance in just two weeks.
Not long after, this representational drift was observed elsewhere as well – in the representation of tones in the auditory cortex, the representation of odors in the olfactory cortex and task representation in the posterior parietal cortex. Representations identified in the brain in any given recording session seemed to break down in a matter of weeks. The conundrum was however, that function remained consistent. Like Wilder Penfield showed 70 years ago, just because the representation of hand movement was no longer where it used to be, doesn’t mean that the person can’t move their hand. The job is being done somewhere else.
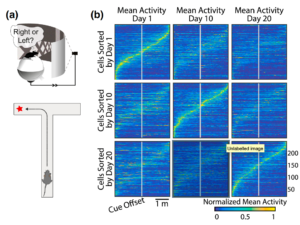
Figure from Rule et al, Current Opinions in Biology (which shows an adapted figure from Driscoll et. al) demonstrates how spatial coding in the post partietal cortex shifts over time. each matrix shows normalized activity of numerous cells such that the same cells with elevated activity on Day 1 are not the same ones as on Day 10 or Day 20.
Yet, how this works is a mystery. Why do things move around? How does the brain ensure that even though the participating neurons are changing, that the job is still done without any loss of function? Is there a benefit to this flux?
One thing is clear. The flux is not totally random. A recent study by Rule et al shows that the drift is not simply a random walk but systematically constrained far above chance so there is some method behind it. Rule et al propose some kind of error representation mechanism in the brain. However, it may well be worth considering the possibility that what if it is not which neuron is responding that even matters at all? For the sake of analogy, you can get your same coffee at Starbucks every day even though the barista changes often – or even when you go to a different Starbucks.